
Abstract
As the Army Malaria Institute entered its fifth decade, its research mission expanded and matured. Five research departments were engaged in assessing a variety of malaria drugs, molecular biology, field, clinical and diagnostic studies while arbovirus vaccines and molecular epidemiology topics were studied. Internal and external reviews of the Army Malaria Institute (AMI) were conducted indicating that AMI should remain within the Joint Health Command and eventually change its name to better reflect its role within the entire Australian Defence Force and with infectious diseases beyond malaria. AMI’s deployment capability is intended to be emphasised by the evolution of a separate identifiable unit involving the uniformed members. How AMI should manage its quasi-academic status as well as external research funds has not been determined yet. As AMI’s Fiftieth Anniversary approaches in mid-2016, it is clear that the on-going threat of infectious diseases to the ADF will mean that the Institute will continue to evolve its structure and functions into the future.
Mission, Organisation and Staff
As the ADF’s overseas missions have changed in the new millennium with a more global involvement of the Defence Organisation, so the role of AMI has evolved to match the infectious disease threats encountered. Once the malaria problems on East Timor were largely solved by stabilisation of the civilian population and improved compliance with standard malaria chemoprophylaxis by ADF personnel, little malaria has occurred in Australian soldiers.1 malarial infections in the ADF peaked in 2000 and since then have been at a relatively low level. There were few malaria infections among the ADF’s contribution to the Regional Assistance Mission to Solomon Islands (RAMSI) from 2003-2013 despite being deployed in a malaria endemic area. Iraq, other than possibly parts of Kurdistan, is free of malaria, whereas the exposure risk during operations in Afghanistan was largely limited seasonally to mid-year in irrigated valleys and areas near the border with Pakistan. Although there were reports of a few soldiers in the SAS Regiment being infected with falciparum malaria while in Afghanistan, AMI was only able to confirm these findings in three cases from the limited clinical material received in Australia. Similar scattered reports of malaria in ADF observers in Southern Sudan also have been received but not confirmed with the blood specimens available to AMI.
The resumption of joint military exercises in Papua New Guinea in 2014 and 2015 (e.g. OP Olgetta Warrior, a company-sized exercise on the northern coast of PNG) has resulted in up to 5% of the ADF soldiers deployed developing vivax malaria on return to Australia 1-5 months post-exercise. Although some cases may be due to sub-optimal compliance with post-deployment primaquine eradication courses, it also is thought that some of the prophylaxis failures were due to cytochrome polymorphisms resulting in the poor metabolism of primaquine.2 Although few ADF members travel to Africa, those that do may be exposed in high risk areas for falciparum malaria.
AMI has confirmed that the infection of one soldier with falciparum became infected whilst on a safari holiday in Tanzania. In June 2015, four Australian sailors participating in OP Manitou in the Indian Ocean went on leave to a Tanzanian beach resort. Two weeks later while aboard HMAS Newcastle in the Indian Ocean they developed acute febrile illnesses. Despite being unable to formally confirm the diagnosis of falciparum malaria due to a lack of diagnostic capability aboard, quick clinical action by the ship’s medical officer in consultation with AMI resulted in a therapeutic response and recovery of all four falciparum cases (later confirmed by molecular methods) using atovaquone/proguanil treatment.3
The ADF experienced more cases of dengue in East Timor than in any other deployment since the Second World War. After an initial surge of several hundred cases in the first years of the deployment4, a small number of cases continued to be diagnosed until the engagement ceased in 2013. Monitoring of blood specimens collected from febrile soldiers confirmed infections with all four dengue virus (DENV) serotypes. Dengue cases also were diagnosed routinely among ADF personnel deployed as part of the Regional Assistance Mission to the Solomon Islands (RAMSI). Although ADF members in northern Queensland and the Northern Territory are operating in areas of high Ross River and Barmah Forest virus transmission, few acute infections have been confirmed.
The end of major military operations in Iraq and Afghanistan along with governmental fiscal limitations triggered by the Global Financial Crisis of 2008 lead to a re-evaluation of the ADF’s roles and missions, which is an on-going process through a series of White Papers, First Principals’ and Force Structure Reviews. As with the larger military, AMI was also subjected to both internal and external reviews to determine if the Institute had the appropriate mission, resources and capability, particularly in terms of AMI’s overseas operations. This process was triggered by AMI’s use of external funds from various private and public sources and the need for the ADF to be certain that these monies were being handled appropriately. The external review chaired by COL Craig Schramm reported in 2012 and the major findings included the confirming the great value of AMI to the ADF, the need to improve communication between AMI in Brisbane and Joint Health Command (JHC) in Canberra, that AMI should remain as a part of JHC and that a unit name change was needed to better reflect the broader function of AMI. Furthermore the review found that AMI was under-resourced for its activities such that subsequent external funding should be approved by JHC under an as yet to be determined mechanism.4 A further internal review reporting in 2014 was conducted within JHC in order to determine how such recommendations might be executed. Recommendations of the internal review included the need to formalise AMI’s relationship with the University of Queensland’s School of Population Health as well as the QIMR Berghofer Medical Research Institute and further development of a process to utilise external funds within the Commonwealth financial process.5
Although several options for the re-organisation of AMI were considered during the reviews, it was felt that there was little scope to change the mixed APS / military workforce while maintaining the core mission and capability of the unit. Of particular concern were AMI’s overseas operations such as malaria surveys in Melanesia and surveillance for arboviral diseases in the Pacific as they occurred within the civilian scientific community and outside any other ADF structure. The resolution of AMI’s unique situation of being a military unit working largely within a civilian scientific community was a continuing process through to the time of the Institute’s 50th anniversary on 16 Jun 2016.
A major staff change occurred when after twenty years of leading AMI, Prof Karl Rieckmann retired and Dennis Shanks became the second AMI Director in 2006. This transition maintained a great deal of continuity as Prof Shanks had previously been assigned as a US Army exchange officer at the Army Malaria Research Unit in Ingleburn from 1992-94 and was well-known to the longer serving staff members. The military leadership of AMI changed from LTCOL Bob Cooper to LTCOL Ivor Harris in 2011 and then subsequently to LTCOL Alyson Auliff in 2014; the latter two officers being designated as the Senior Scientific Officer/Administrative Commanding Officer rather than a Commanding Officer.
Scientific achievements and accomplishments at AMI over the last decade to 2015 are reviewed by research department: Drug Resistance and Diagnostics, Clinical Studies and Surveillance, Drug Evaluation, Vector Surveillance and Control, and Arbovirology.
Drug Resistance and Diagnostics
The mission of the department of Drug Resistance and Diagnostics (DRD) is to improve malaria diagnosis and inform treatment policy in order to reduce ADF non-battle casualties. Early diagnosis and treatment are critical to prevent severe complications and death caused by malaria parasites, particularly in non-immune persons such as nearly all ADF personnel. Accurate and rapid diagnosis of malaria is often challenging in remote areas and in areas with low transmission. The development and spread of drug resistance in malaria parasites has also become a major obstacle for the effective treatment and control of malaria globally. Understanding the mechanism and speed with which parasites develop drug resistance enables one to identify resistance markers and use these markers to detect and monitor the spread of drug resistant parasites. The outcomes inform treatment policy to ensure effective drug regimens are used to treat resistant parasite infections and help to develop new drugs to overcome resistance. Therefore, DRD’s focus over the past decade has been to improve malaria diagnosis and to understand why and where antimalarial drugs are ineffective due to resistance.
DRD personnel consist of two APS and two Army research scientist positions. Dr Qin Cheng was the Head of DRD over the entire decade. The military positions have changed over time. After completing a PhD in 2012 (supervisors: Q Cheng, M O’Neil, D Gardener) Major Alyson Auliff was promoted to LTCOL and Senior Scientific Officer of AMI in 2014. In the same year Major Joanne Baker was posted out of AMI following her completion of a PhD in 2011 (supervisors: Q Cheng and JS McCarthy). LT Simone Dowd and CAPT Chris Peatey joined DRD in 2012 and 2013, respectively. LT Michael Korsinczky, a Reserve Officer, became inactive at the end of 2008 after 10 years service at AMI. Three other reserve officers also contributed to DRD’s activities: CAPT Darren Krause (2006 – 2010), CAPT John Hymus (2006-2007) and CAPT Josh Mylne (2010 – 2012). DRD also has had a varying number of civilian scientists funded by national and international grants. During this decade, Dr Cheng successfully obtained two NIH RO1 grants (2004-2007 and 2007- 2010, in collaboration with Prof Dennis Kyle), a NIH R21 grant (2012-2014, in collaboration with Prof John Adams), a NHMRC project grant (2012-2015) and a GEIS grant (2011-2015, in collaboration with LTC Norm Waters). These grants, administrated by QIMR Berghofer Medical Research Institute, funded Mrs Jenny Peters (2004-2007), Dr Darren Krause (2004 – 2007), Dr Franka Teuscher (2008 – 2010), Ms Karen Gray (2012 – 2014), Ms Karryn Gresty (2011 – 2015), Dr Chris Peatey (2012), Mrs Karen Anderson (2013 – 2015) and Dr Cielo Pasay (2013 – 2015) conducting research and surveillance activities in DRD. AMI also received AusAID funding through Pacific Malaria Initiative Supporting Centre, University of Queensland (2009 – 2012) which supported Ms Lisa Bain (2009 – 2011), Mr Wesley Sharrock (2009 – 2010), Ms Karen Gray (2009 – 2011) and Ms Simone Dowd (2010 – 2012) to perform epidemiological studies and surveillance as part of the DRD team.
Improving malaria diagnosis: DRD has been actively exploring and evaluating novel malaria diagnostics which include molecular and rapid diagnostic tests (RDTs) most of which consist of immunochromatographic devices. Malaria RDTs offer an alternative to microscopy for accurate and rapid diagnosis of malaria infection particularly in remote areas, but their performance is highly variable. DRD is a partner in the WHO-FIND Malaria RDT
evaluation Network, having participated in product testing of over 200 commercially available malaria RDTs. The results of these testings enabled products to be ranked based on their performance and produced a list of quality malaria RDTs on which to base the procurement decisions of the WHO and other UN agencies, NGOs, national malaria programs and the ADF. These quality-tested RDTs are a fundamental tool for ADF personnel deployed to remote areas where electricity and microscopists are not available (https://www.WHO.int/RDTs). At DRD’s recommendation, ADF has now changed to best performing RDTs that meet the WHO procurement guideline.
Factors influencing the performance of malaria RDTs including parasite strains, lack of antigen, the amount and type of antigens6-9, antibodies used in RDTs10-12 and competitive human antibodies13 were investigated in collaboration with Prof James McCarthy. These activities were partially funded by the WHO & FIND. One important discovery was that a high proportion of P. falciparum isolates in Peru lack a major target antigen (HRP2) and thus are not detectable by most RDTs.14 This finding led the WHO to recommend the use of an alternative diagnostic strategy in affected areas and led to large scale surveillance on the prevalence and geographic distribution of these parasites globally.15 The amount of antigen and competitive human antibodies used were also found to significantly affect the sensitivity of RDTs.9,13,15,16 This research has helped to improve the design of RDTs and provided important information about suitable diagnostic tools for use in South America. Besides RDTs, DRD has also been exploring new molecular tools for detecting malaria parasites in collaboration with Prof James McCarthy at QIMR Berghofer Medical Research Institute.
Confirmation of malaria diagnosis for ADF personnel:
DRD staff performed molecular diagnostic tests (PCR) of samples from ADF members suspected of having malaria infections, to verify the initial diagnosis and to confirm parasite species. This information assists clinicians to better manage patients and to ensure the accuracy of data entered into the ADF central malaria register. DRD also provides molecular confirmation of malaria infections for other organisations on request. In collaboration with colleagues in Westmead Hospital, DRD diagnosed the first case of P. knowlesi infection in Australia from a traveller.17
Monitoring malaria drug resistance in the Pacific region:
Therapeutic efficacy studies of antimalarial drugs in the Pacific region were partially funded by Global Emerging Infectious Diseases and Surveillance (GEIS) of the US Department of Defense). Chloroquine (CQ), alone or in combination with sulphadoxinepyrimethamine (SP), was widely used for the treatment of both falciparum and vivax malaria for several decades in the South Pacific countries of Solomon Islands, Vanuatu and Timor Leste (formerly known as East Timor). In 2008 due to increasing drug resistance, artemisinin combination therapies were introduced in these countries as the first line therapy for malaria. It is important to monitor the clinical efficacy of such drug combinations in these countries. DRD has collaborated with the national Ministries of Health (MOH) and the WHO to assess artemisinin-lumefantrine efficacy in Solomon Islands (2011), Vanuatu (2012) and Timor Leste (2012, 2013). The artemisinin combination was found efficacious in these countries. The outcomes inform treatment policy for both MOH and ADF as this combination is recommended first line therapy for these countries as well as for ADF.
Studies concerning the molecular assessment of parasite drug resistance in the Pacific region have been partially funded by US Defence Department’s GEIS. In collaboration with the WHO and GEIS, DRD has investigated the malaria drug resistance profiles in Solomon Islands and Vanuatu at the time of introduction of artemisinin combination therapies. DRD found that falciparum parasites in both countries had high and moderate levels of
resistance to CQ16 and SP18, respectively, sufficient to cause treatment failures. These outcomes provide baseline data for future drug resistance monitoring and provide health intelligence to GEIS and ADF.
Identifying Drug resistance markers:
Over the past 10 years, the DRD team, led by Dr Qin Cheng, has conducted innovative research into the mechanisms and phenotypes of resistance to artemisinin derivatives in P. falciparum, and CQ, antifolate drugs in P. vivax. The outcomes established drug resistance markers enabling DRD to assess the prevalence and evolution of drug resistant parasites in Asia-Pacific regions such as Timor Leste and PNG where ADF personnel have been and are likely to be deployed.
Studies on artemisinin (ART) inducted dormancy and ART resistance in P. falciparum were partially funded by a US NIH grant in collaboration with Prof Dennis Kyle, University of South Florida (USF). ART combination therapies are now used in all malaria areas as the first-line therapy. However, ART monotherapy with frequent treatment failures and parasites resistant to ART have been reported in several Southeast Asian countries. The underlying mechanisms of treatment failure and artemisinin resistance are not clear. DRD demonstrated that parasites became dormant following treatment with ART, but recover to resume normal growth days and weeks later 19, 20 DRD also identified that the dormancy profile changes as ART resistance develops.21, 22 Because of this research, it is now known why treatment failures occur and how to reduce them. New drug development now includes testing for dormancy induction, and field teams have begun to use the dormancy assay to detect and monitor ART resistance. DRD has since discovered that these dormant parasites are not metabolically quiescent, as the metabolic pathways in apicoplast and mitochondria are active.23, 24 These findings enabled DRD to develop a method to identify quiescent parasites and to use drugs to interrupt their recovery from dormancy.23, 24 DRD continues to investigate the mechanisms of dormancy to discover new means of disrupting this evolutionary adaptation.
Considerable effort has also been invested into identifying molecular markers for ART resistance. The heavy reliance of ART combination therapies for treatment of malaria globally makes this work key to the continued ability of the ADF and others to adequately treat personnel suffering from malaria. In a laboratory-selected drug resistant parasite line, DRD identified a correlation between a resistant phenotype and parasite multidrug resistant genes.25, 26 Further work is underway to find other genetic markers of drug resistance.
P. vivax has developed resistance to the antifolate drugs. DRD identified a unique set of genetic mutations in the parasite that correlated with drug treatment failures.27-29 In collaboration with Prof John Adams at USF, and LTCOL Mike O’Neil at WRAIR, DRD then used advanced molecular technologies to elucidate the impact of these mutations on the drug efficacy. This investigation demonstrated that while some mutations directly interfere with drug binding, other mutations compensate the deleterious effect of these mutations.30-32 These systems validated the mutations as molecular markers for resistance surveillance and provided biological platforms to study the function of P. vivax genes.
Chloroquine and mefloquine resistance in P. vivax was studied in collaboration with Menzies School of Health Research in Darwin NT. The collaborative team demonstrated sequence and copy number polymorphisms in the multiple drug resistance gene 1 in P. vivax correlating with CQ and mefloquine resistance phenotypes in P. vivax.33-35 These changes are being validated as molecular markers for resistance. In collaboration with the P. vivax Genome Project DRD investigated and compared drug interaction genes between P. vivax and P. falciparum contributing to the understanding of the P. vivax genome and differential drug action between P. vivax and P. falciparum.36 This contribution was recognised in a publication in the journal Nature.36
Investigating P. vivax relapses in ADF:
Relapsing P vivax infections is one of the major health problems for ADF, and can occur for several years after the return from a malaria endemic area. DRD has investigated the relapse patterns and intervals as well as the genetics of parasites causing relapses in ADF personnel returning from Timor Leste. DRD discovered that relapses in ADF personnel resulted from clonal activation of hypnozoites (residual hepatic parasites) at predetermined intervals and that multiple strain infections increased the risk of relapse.37 This outcome highlights the importance of taking measures to minimise exposure to mosquito bites. Such information also helps elucidate P. vivax relapse biology in order to reduce the risk of relapse and to develop better drugs against P. vivax. Besides parasite factors, DRD has also begun investigations of host factors contributing to malaria relapse.
Investigating molecular epidemiology of malaria in the South Pacific region:
DRD was part of the Australian Government Pacific Malaria Initiative team assisting the malaria elimination efforts in the Solomon Islands and Vanuatu. This project was funded by AusAID through the University of Queensland. Using molecular tools, DRD has determined malaria prevalence, the type of parasites, as well as mapping their distribution 38-41 and drug resistance profile.16, 18 DRD also identified several challenges to diagnostics in elimination settings.41,42 These data provided important epidemiological information on the current malaria status as a baseline to measure progress against, a guide for targeting elimination efforts in the country and valuable health intelligence to the ADF. These activities supported the whole-of-government strategy, and demonstrated ADF’s contribution to global health. AMI demonstrated its capability for malaria risk assessment for future military deployments. DRD also conducted and was involved in other studies.43-56 In summary, the research activities conducted within the DRD department helped to ensure that wherever ADF personnel are deployed, good diagnostic tests and appropriate drug treatments are provided.
Clinical Studies and Surveillance
The Department of Clinical Studies and Surveillance (CSS) as the translational medicine component of AMI has conducted several clinical trials over the last decade. CSS has been successively lead by LTCOL Peter Nasveld, MAJ Andrew Ebringer and LTCOL Ken Lilley. These studies have included a dengue vaccine study 2010-12 in Australia using a quadravalent live-attenuated dengue vaccine to determine safety and immunogenicity while parallel efficacy studies in SE Asia were completed by other investigators.57 A primaquine short course (1 week instead of 2) study was conducted in Timor Leste in 2009 to determine if a modified post-exposure regimen of primaquine was tolerated and safe in returning ADF soldiers.58 Multiple malaria surveys were completed in Vanuatu and Solomon Islands during 2008-12 including Tanna and Erromango Islands in Vanuatu as well as Santa Cruz, Isabel and Nggella Islands in Solomon Islands.41 CSS participated in a health survey of Papua New Guinea (PNG) Defence Force personnel during 2008 examining otherwise healthy soldiers for malaria, tuberculosis and other infectious diseases in both Port Moresby and Lae. Vaccine studies performed at AMI included a Japanese Encephalitis (JE) study during 2003-06 which was a randomised double-blind evaluation of safety and immunogenicity of a genetically modified JE vaccine starting from a yellow fever vaccine virus backbone.59 A 5-year follow up of the same JE vaccine during 2005-11 was undertaken in volunteers in order to determine the booster requirement for those deploying repeatedly into endemic areas of SE Asia.60 This JE vaccine is now registered by the Therapeutic Goods Administration (TGA) in Australia and used by the ADF as a prime example of how AMI studies allow the ADF to access new and improved vaccines.
CSS also studied the use of the long-acting primaquine analogue, tafenoquine, which was tested for the treatment and prevention of relapse in soldiers returning from Melanesia.61-63 In addition, three different post-exposure tafenoquine treatment regimens were used to treat vivax malaria relapses in Australian soldiers following deployment to Bougainville PNG and East Timor.64 CSS has also conducted WHO Certification courses in malaria microscopy for ADF Scientific Officers and selected allied and civilian personnel. CSS provides an external quality assurance program in malaria microscopy for ADF Scientific Officers to facilitate their maintenance of skills and to prepare them for diagnostic testing during deployment.
Drug Evaluation
The mission of the Drug Evaluation (DE) Department is to provide effective and well tolerated antimalarial drugs for the protection and treatment of ADF personnel against malaria infections. In order to provide the ADF with the most effective antimalarial drugs, DE conducts preclinical evaluation of new candidate antimalarials and participates in the optimisation of drug regimens in clinical studies. The antimalarial drug discovery program in DE involves parasitological, pharmacological and animal studies with compounds synthesised by medicinal chemists from academia or the pharmaceutical industry. For a candidate compound to advance in preclinical development, an established series of investigations are pursued. The compound must possess good in vitro antimalarial activity against P. falciparum and P. berghei lines and is not highly cytotoxic in mammalian cell lines. Compounds that show high in vitro potency and low toxicity are then assessed in mice for tolerability and in vivo efficacy against rodent malaria. The most promising compounds are further evaluated for efficacy and safety at AMI in Aotus monkeys infected with the chloroquine-resistant FVO strain of P. falciparum or the chloroquine- and antifolate-resistant AMRU1 strain of P. vivax. Additional studies including stage of action against parasites, mechanism of action and pharmacokinetic properties (i.e. blood concentration-time profile) are pursued with the lead compounds. Those compounds that successfully complete the paradigm of research, including toxicity studies that are performed external to AMI, are then considered for first-time in-human studies which are done with collaborators at QIMR. DE’s role in clinical investigations is to assist in the conduct of efficacy studies of antimalarials and to perform pharmacokinetic and ex vivo antimalarial studies to evaluate the pharmacokinetic-pharmacodynamic relationship of standard antimalarials, so ADF personnel receive safe and effective drugs, particularly for the treatment of multidrug-resistant falciparum malaria.
Dr Michael Edstein was the Head of the department during the entire decade. Members of his staff included Dr Barbara Kotecka (until 2008), Dr Marina Chavchich (from 2003), Dr Geoffrey Birrell (from 2008), Mr Thomas Travers, Ms Donna MacKenzie (from 2010) Sergeant Kerryn Rowcliffe, Sergeant Scott Smith (until 2012) and Lieutenant Henry Simila (from 2011). Veterinarians at the Institute’s animal facility were: Lieutenant Colonel Ivor Harris, Captain (Army Reserve) Joanne Beckett (from 2002), Captain (Army Reserve) John Hunter (from 2009) and Captain Fiona McCallum (from 2012). Animal technicians included WO2 Stephen McLeod- Robertson (from 2005), Mr Zbigniew Kotecki (until 2008), Ms Nerissa Walpole (until 2013) and Anthony Kent (from 2014).
Dr Edstein was a co-investigator on two NHMRC (2012-2015) grants and a Medicine for Malaria Venture grant (2007-2009). The NHMRC grant with Eskitis Institute for Cell and Molecular Therapies (Griffith University) and sponsorship from Jacobus Pharmaceutical Company co-funded Mrs Karin van Breda (2012-2015) for preclinical drug investigations at DE.
Antimalarial drug discovery program:
Activities and achievements of the Department included collaboration with several university groups (i.e. Eskitis Institute for Cell and Molecular Therapies at Griffith University; School of Chemistry and Molecular Biosciences at The University of Queensland; Monash Institute of Pharmaceutical Sciences at Monash University; School of Physical, Environmental and Mathematical Sciences at the Australian Defence Force Academy, Faculty of
Health Sciences at North-West University-South Africa), the Land Division at the Defence Science Technology Organisation, the Novartis Institute of Tropical Diseases and Jacobus Pharmaceutical Company in the preclinical development of new antimalarial compounds. These collaborators synthesised compounds for evaluation at DE with unique chemical structures and modes of action. The diverse chemical structures included second-generation artemisinin derivatives65, a third-generation triazine66, phenolic glycosides67, thiaplakortones68,69, pyrroloiminoquinones70, acyclic nucleoside phosphonates70-74, dinuclear rutheniums75, and aminomethylphenols.76
In general, these compounds possessed 50% in vitro inhibitory concentrations (IC50s) <1 μM against P. falciparum lines and low cytotoxicity in mammalian cell lines with selectivity index (IC50 of cell line / IC50 of P. falciparum line) usually >50. The most promising compounds advanced to mouse studies. Of these compounds, several were found to be well tolerated in mice at a dosage up to 64 mg/kg/day for 4 days with a low ED50 (50% effective dose) value of <2 mg/kg/day in suppressing blood schizonticidal activity in the murine-P. berghei Peters 4-day test.
So far the most promising class of compounds identified at AMI are the aminomethylphenols, with JPC-2997 and its analogues being highly active in vitro against the multidrug-resistant P. falciparum lines, with IC50s < 34 nM and remarkably low cytotoxicity (IC50s >35 μM) in mammalian cell lines.76 JPC-2997 possesses potent in vivo suppression activity against P. berghei with an ED50 of 0.5 mg/kg/day following oral dosing in the Peters 4-day test. The radical curative dose of JPC-2997 was remarkably low at a total dose of 24 mg/kg using the modified Thompson test. JPC-2997 was effective in curing Aotus monkeys infected with AMRU1 P. vivax strain at a dose of 20 mg/kg daily for 3 days. Preliminary studies of JPC-2997 in mice show linear pharmacokinetics over the range 2.5 to 40 mg/kg, wide distribution to tissues and an elimination half life of 49.8 hours. The high in vivo potency data and lengthy elimination half-life of JPC-2997 suggests that it is worthy of further preclinical assessment as a partner drug for malaria treatment or as a potential prophylactic agent.
Efficacy of artemisinin based combination therapy (ACT):
In 2001, the World Health Organization recommended ACTs for the treatment of uncomplicated P. falciparum malaria. The ADF first-line treatment for P. falciparum and P. vivax malaria is the fixed ACT of artemetherlumefantrine (Coartem®). Since 2006, there has been increasing reports of reduced susceptibility to ACTs in Southeast Asia with confirmed artemisinin resistance in Cambodia, Thailand, Myanmar and Vietnam. Until new antimalarial drugs are developed that do not require an artemisinin as part of a drug combination, the efficacy of existing ACTs need to be monitored, especially the development of drug resistance.
Under the auspices of the defence cooperation between the Vietnam People’s Army (VPA) and the ADF (2000-2012), AMI in collaboration with the VPA Military Medicine Department carried out a series of clinical studies in south-central Vietnam to compare the efficacy of various ACTs (artemisinin-piperaquine, dihydroartemisinin-piperaquine, artesunateamodiaquine and artesunate-azithromycin) for the treatment of uncomplicated falciparum malaria.76-79
These studies were conducted in an ethnic minority community at Phuoc Chien Commune, Ninh Thuan Province between 2006 and 2010. The ACTs were well tolerated and highly efficacious, with a PCRcorrected cure rate of >96% for the four ACTs. These findings suggest that the population of P. falciparum parasites at the field site were still susceptible to ACTs.
Prior to conducting the efficacy studies in the field, pharmacokinetic and ex vivo antimalarial studies in malaria patients and healthy volunteers were pursued at two military hospitals in Vietnam. These studies demonstrated the bioequivalence of two formulations of dihydroartemisinin-piperaquine 80and the pharmacokinetic- pharmacodynamic relationship of the partner drugs of the two ACTs, dihydroartemisinin-piperaquine 81 and artesunateazithromycin. 82 The artemisinins were responsible for most of the ex vivo antimalarial activity, with a delayed contribution by the partner drugs, piperaquine and azithromycin.
In 2012, DE and US Navy Medical Research Unit 2 (later US Naval Medical Research Center-Asia) successfully obtained US Department of Defence GEIS funding to collaborate with the VPA in the identification and monitoring of the spread of drug resistant malaria in Vietnam.
Dose optimisation of primaquine for postexposure prophylaxis:
Deployment of ADF personnel on peace-keeping duties in Bougainville (PNG) and Timor Leste has highlighted the challenge of preventing P. vivax infections. Primaquine is the only drug commercially available for both post-exposure prophylaxis and radical cure of vivax malaria. A limitation of standard primaquine treatment is that it has to be given daily for 14 days to minimise gastrointestinal disturbances, which is a cumbersome regime contributing to poor compliance. To address this limitation, a primaquine regime of twice daily administration of 30 mg for 7 days was evaluated in 203 ADF personnel returning from peace-keeping duties in Timor Leste.58 The regime was found to be safe and well-tolerated in most subjects. However, with 5% of participants experiencing peripheral cyanosis it appears that this may be the maximum acceptable short-course primaquine dosage in a predominately Caucasian population for the treatment and post-exposure prophylaxis of P. vivax malaria.
In addition to optimising the primaquine regime, we showed that healthy female Vietnamese soldiers had significantly lower clearance of primaquine compared with males, resulting in higher plasma concentrations and exposure of the drug.83 These findings suggest that females may be at greater risk of toxicity with primaquine when administered at the same maintenance dose as males. Larger studies in different ethnic groups are warranted to determine whether a dose adjustment is required for females administered primaquine to minimise adverse events and maintain full effectiveness.
Other studies involving the department during this decade included characterisation of the in vitro susceptibility of a P. falciparum line for future human challenge studies, the in vitro response of P. vivax field isolates to chloroquine84, comparison of various primaquine and tafenoquine regimens for post-exposure prophylaxis to vivax malaria64, population pharmacokinetics of halofantrine for the treatment of falciparum malaria85 and worldwide pooled analysis of parasite clearance half-lives of artemisinin derivatives and ACTs for the treatment of uncomplicated P. falciparum.86
Vector Surveillance and Control
Vector biology is critical to the understanding of how pathogenic organisms spread though insects. The Department of Vector Surveillance and Control (VSC) at AMI has conducted malaria vector studies of the distribution and transmission potential of mosquitoes in Vietnam (2002-2010), Vanuatu (2007) and Solomon Islands (2008-2012).87-89 In addition, VSC has provided entomological support to 2 General Health Battalion’s Health Assessment Team for surveillance of vectors and arboviruses at the Townsville Field Training Area (2010), Shoalwater Bay Training Area (2011), Bradshaw Training Area (2013) and Cowley Beach Training Area (2014).90
Repellents have been studied by comparative tests of the standard ADF insect repellent formulation against a variety of comparable products both commercial and military.91-98 Field studies showed that a commercial formulation of deet was as effective as the ADF formulation, and it was recommended for use by the ADF (Frances et al 2010, Frances et al 2014 and Frances 2013, 2015). Much work has been done with permethrin treatment of ADF disruptive pattern combat uniforms (DPCU) where AMI has conducted an initial evaluation and then recommended the treatment and retreatment regime of DPCU and ADF mosquito bed nets.95, 99, 100 Other investigations have been conducted into rickettsial diseases such as scrub typhus.101, 102; 103 In addition VSC has annually conducted a vector borne disease surveillance and control course for initial employment training for ADF Preventive Medicine Technicians.
Malaria vectors in Papua New Guinea:
The vectorial status of the anopheline species identified in Papua New Guinea during Operation Anopheles was determined using an enzyme linked immunosorbent assay and species specific monoclonal antibodies for the sporozoite stage of the human malaria species. The studies identified 11 anopheline species as vectors of malaria. Of these, the major vectors, based on their distribution, abundance, association with humans, and ability to develop the parasite, were all members of the Anopheles punctulatus Group: Anopheles farauti, Anopheles farauti 4, An. hinesorum, An. koliensis, and An. punctulatus.88 Consolidating the results of the field surveys conducted under Operation Anopheles and the molecular analysis of the material collected, two reviews were published which update our current knowledge of the evolution, distribution, biology, and control of these malaria vectors.104, 105
Malaria vectors of the Solomon Islands: Knowledge of a vectors biology and behaviour is critical to understanding how vector borne diseases are transmitted and how vectors might be controlled. In the Solomon Islands the major malaria vector is Anopheles farauti. Observations on this species, following the failure of malaria eradication programmes in the early 1970s, showed that An. farauti, under insecticidal pressure, began feeding early in the evening (6pm – 9pm) and predominately outdoors.106 This allowed for avoidance of the insecticide inside the houses and the continuation of malaria transmission.107 This type of biting behaviour will also compromise the effectiveness of the currently in use vector control strategy of insecticide treated bed nets (ITN) which also relies on the vector entering the house when the occupant is asleep under the ITN.108 With the support of PacMISC, studies in Temotu and Santa Isabel Provinces confirmed that this early outdoor biting behaviour is still maintained in the An. farauti populations of these two provinces.109-111 This observation is of concern as this will make the desired goal of malaria elimination difficult to achieve with ITN alone. The studies in Temotu Province showed that the only anopheline present was An. farauti, however in Santa Isabel Province several species have previously been recorded, though these were from surveys conducted over four decades ago.112 These early surveys identified An. punctulatus and An. koliensis, both very efficient vectors of malaria, as occurring in Santa Isabel Province. The recent surveys failed to find either of these species, but An. farauti, An. hinesorum and the three members of the Anopheles lungae complex: Anopheles lungae, Anopheles solomonis and Anopheles nataliae were found to be common and wide-spread. The members of the Anopheles lungae complex are not considered vectors of malaria as they rarely bite humans, however during these recent studies conducted in Santa Isabel An. solomonis was the dominant anopheline biting humans in an inland village.109
Further studies on the bionomics of An. farauti on the north coast of Guadalcanal demonstrated that this vector prefers to breed in brackish pools formed by creeks blocked to the sea by sandbars; these conditions occur most frequently at times of low rainfall.110, 113 Thus, unlike most other malaria vectors in the region, An. farauti is most abundant at the end of the dry season (October – December). This information is important in determining the malaria transmission season and in implementing control strategies.
The avidity of a mosquito species for human blood is an important parameter in gauging a mosquito species efficacy as a malaria vector. However determining the host preference of a species is difficult in the field since to obtain an unbiased sample sufficient numbers of blood fed specimens must be collected by searching for them resting outdoor in the surrounding jungle. This method is laborious and not particularly productive, thus this
parameter of a vector’s transmission capacity is often poorly calculated. To improve on this method the use of portable fences (shade cloth: 2 m high x 20 m long) were trialled. These were erected in villages between the available blood sources (predominantly human, pig, and dog) and the main breeding sites. It was found that the fences effectively intercepted An. farauti post feeding, allowing for an unbiased sample to be collected in sufficient numbers.114
In the early 1990s insecticide treated nets were introduced into the Solomon Islands and Vanuatu as the principal method of malaria control, largely replacing indoor residual spraying. These nets needed regular retreatment to be effective. In 2008 these nets were superseded by long lasting insecticidal nets (LLIN) which were reported to be effective for 3-5 years without retreatment. In Vanuatu the efficacy of LLIN was evaluated as to the condition of the net and its ability to kill mosquitoes after 2 and 3 years of use in rural village communities. It was found that the condition of the nets (number of tears and holes) and the insecticidal properties of these nets began to deteriorate after 2 years with the conclusion that the nets should be replaced with new nets every three years.115
Arbovirology
The Arbovirology Department has focussed on measures to prevent mosquito-borne viral (arboviral) diseases in the ADF and mitigating the consequences of infections that cannot be prevented. AMI was fortunate to have a high level biosafety Physical Containment Level III laboratory established in 2006. This was a significant development, after several years of preparation, and provided the ADF with the capacity to diagnose arboviral infections in service personnel returning to Australia rather than hearing about these events second hand. The capacity to genotype arboviruses also gave the ADF a capacity to investigate claims that returning personnel were responsible for dengue in Australia. Ten years later, AMI and the ADF will have to consider how to maintain this important capability despite the turnover in staff competent to use this facility and limitations associated with training new staff members.
The trial of the Chimerivax™ (Sanofi Pasteur) vaccine against Japanese encephalitis was initiated by the Arbovirology Department in response to the incursion of Japanese encephalitis virus into northern Australia. There also was dissatisfaction with the inactivated vaccine that was being used for personnel deployed to Timor Leste and other south-east Asian localities.59, 60 The old Japanese encephalitis vaccine was produced in the brains of
mice and required three injections over a period of 3-6 months to elicit protection and thus was not useful in a rapid-deployment scenario. This clinical trial faced enormous hurdles being the first live, genetically
modified, exotic human pathogen for intentional use in Australia. LT/CAPT Mark Reid spent almost two years negotiating with various regulatory agencies, often with conflicting legislation and regulations, to gain approval for the vaccine trial to go ahead. This included experiments to demonstrate that local mosquitoes could not become infected if they fed on viraemic vaccinees.116 As a result of these trials, Australia became the first country to license this vaccine (IMOJEV™, Sanofi Pasteur) and ADF personnel now can be deployed within a few weeks of having a single dose of this vaccine knowing it confers protection on 80% of recipients for up to 5 years. Two doses of vaccine confer 90% protection at this interval.
The Arbovirology Department also provided laboratory support to the phase I/II trial of a killed Ross River virus vaccine which showed the vaccine to be safe and determined the dose and formulation of vaccine for use in the phase III trials which have just been completed.117 Ross River virus is believed to have been responsible for epidemics of polyarthritis among garrisoned troops in northern Australia in World War II. While Ross River virus has only been a health issue in Australia and the Pacific, foreign military personnel participating in field exercises in Australia have developed clinical infections with this and other Australian arboviruses. The mosquitoes found in Singapore and the southern states of the USA are competent to transmit Ross River virus if it were to be introduced by returning military personnel which suggests that the Ross River virus vaccine may have utility as an adjunct to biosecurity precautions.
Much of the basic research undertaken by the Arbovirology Department has focussed on understanding the population dynamics of regional arboviruses in order to inform vaccination strategies, where vaccines are being developed, or to aid the development of more effective mosquito control programs.118-122 The Arbovirology Department also is attempting to identify the epidemiological risk factors that determine why outbreaks of dengue occur when and where they do. Many of the viruses required for this research have been obtained from the diagnostic service AMI provides for the ADF as well as from support provided for the dengue surveillance
efforts of smaller Pacific Island nation states and in developing countries in south-east Asia.123-125 In 2010, the Arbovirology Department predicted the outbreak of dengue due to DENV 3 that occurred in the Pacific in early 2013.124
The Arbovirology Department has a long-standing collaboration with the US Armed Forces Research Institute of Medical Research in Bangkok focussing, principally, on dengue.126 In 2006, MAJ Aaskov had the opportunity to spend several weeks with the Virology Department at the Walter Reed Army Institute of Research in Washington on a Prince of Wales Fellowship – with a focus on the molecular determinants of outbreaks of dengue and on dengue vaccine development. In 2004, Senator Hill, Minister for Defence, announced that military co-operation with the Vietnam People’s Army would be expanded to include a $1.5 million, six year, dengue project. Initially, this involved establishing a network of laboratories at Military Hospitals through central and southern Vietnam able to undertake dengue surveillance and training staff to perform this role. This complemented the Vietnam Ministry of Health National Dengue Plan which MAJ Aaskov, in his civilian role, had helped prepare. While initially very successful, subsequent changes to the plan diminished the long term impact of the project. Nonetheless, it did demonstrate major weaknesses in many regional communicable disease surveillance systems and the ineffective and wasteful nature of most dengue control measures. On a positive note, one Vietnam People’s Army Officer obtained her MSc through the collaboration. She was able, for the first time, to map the movement of strains of dengue virus throughout a country in significant detail and demonstrate the critical role for viraemic humans and unsophisticated transport systems in the movement of dengue viruses over long distances. There has been a significant transfer of microbiological skills to Vietnamese colleagues and development of an appreciation of quality processes in laboratory diagnoses. Informal scientific collaboration has continued between AMI Virologists and VPA Officers, particularly at the Military Institute of Preventive Medicine, formerly the Military Institute of Hygiene and Epidemiology.
Other Viral Infections
AMI has developed an interest in viral infections other than dengue and Japanese Encephalitis when these infections were likely to stop military operations. Pandemic influenza is one such infection which was studied using historical records from the Australian Imperial Force (AIF) of the First World War, specifically the 1918-19 influenza pandemic. This was a collaborative effort between the Australian Defence Force Academy at the University of New South Wales and the Centre for Military and Veterans Health at the University of Queensland. The main finding has been the highly variable mortality effect of 1918-19 influenza pandemic on Allied military units. Some Australian military units had >1% of their personnel die while other similar units next to them had no deaths. This unexplained heterogeneity gives a means of dissecting the epidemiological causes of mortality during the pandemic especially given the ability to control for other factors in a military environment.127-130 Naval ships had few deaths during the pandemic despite large infection rates unless they were on isolated patrol duty in the Southern Hemisphere where >5% of the crew died on some cruisers.131, 132 Such findings indicate that one’s previous history of respiratory infections particularly in the recent past largely determined one’s mortality risk during the pandemic. Expanding the influenza work has also created collaborations with the two US Military Academies at West Point and Annapolis. Further investigations have involved New Zealand collaborators using New Zealand Imperial Force and civilian data.133-136
The military is interested in what happens when lethal viral infections occur in non-immune, isolated populations such as on small Pacific islands. As part of the influenza work described above, small island populations in the Pacific were examined.137-139 Epidemic viral disease including but not limited to influenza devastated the Pacific Islands in the 19th and early 20th centuries. Population collapse of up to 90% allowed the disappearance or
displacement of many Melanesian, Polynesian and Micronesian cultures. This was further extended to measles infections on Pacific islands and other closed populations which have distinctly different immunological consequences to influenza. Measles was particularly devastating even to adults. Their vulnerability to lethal epidemics was widely observed, but never explained. Using unique data sets from the isolated island of Rotuma, we have described one of these first contact measles epidemics in great detail and are now working on its possible genetic components.140 A historian at the University of Cape Town has given AMI detailed mortality records from the Boer War concentration camps where half of the children died largely of pneumonia and measles.138 Further comparisons of the lethality of measles in the very different settings have been made in the hope of further defining the host’s immune response and mortality risk factors for future epidemics of exotic viruses.
Veterinary Medicine and Animal Facility
The Animal Facility at AMI consists largely of its entirely locally-born Aotus monkeys. The Aotus monkey -P. falciparum/P. vivax model is the major non-human primate model for evaluating the efficacy of new candidate antimalarial compounds. The colony of approximately 60 monkeys (as of 2015) has been raised from animals originally obtained from the Walter Reed Army Institute of Research (WRAIR) in Washington DC. The use of Aotus monkeys as a primate malaria infection model approximating the disease in humans is well established and is their primary use at AMI.141, 142 Mice are also used in an early drug screening method using P. berghei. LTCOL Ivor Harris was the primary veterinary officer for the fifth decade of AMI’s existence with CAPT Fiona McCallum taking over that role from 2014. AMI has a vigilant and effective Animal Ethics Committee which ensures compliance with the National Health and Medical Research Council Australian Code for the Care and Use of Animals for Scientific Purposes.
Conclusion:
The Army Malaria Institute which had its beginnings in the Sinai Desert in 1916 has served to protect Australian soldiers during both World Wars and in multiple regional conflicts since Vietnam when it was formally re-established at the University of Sydney in 1966.127, 143, 144 Over the last fifty years, in its current configuration at Gallipoli Barracks in the Enoggera suburb of Brisbane since 1996, AMI’s mission has expanded to include other infectious diseases capable of stopping military operations, such as dengue. This expanded role beyond malaria now includes all three armed services. As the name change indicates, the Institute will continue to evolve into the future to provide the best possible protection to the entire ADF when deployed into areas of infectious disease risk whether in Australia or across the world.
Staff and facilities
2005 Prof G. Dennis Shanks returns to AMI and becomes second Director of Army Malaria Institute in 2006
2006 MAJ Andrew Ebringer arrives at AMI, CAPT Darren Krause (2006 – 2009) and CAPT John Hymus (2006
– 2007) join AMI as reserve officers MAJ Michael O’Neil arrives as US Army assigned officer
2007 LT Michael Korsinczky becomes inactive from Army Reserve
2008 CAPT Geoffrey Birrell arrives at AMI
2009 LTC Norman Waters arrives as US Army assigned officer
2010 LT Michelle Rourke, LT Fiona McCallum arrive at AMI; CAPT Josh Mylne (2010 – 2012) joins AMI as a
reserve officer
2011 Dr Wen Liu arrives at AMI LTCOL Bob Cooper transitions to Army Reserve
2012 CAPTs Weng Chow, Simone Dowd, and Chris Peatey as well as LT Lisa Rigby arrive at AMI LTCOL Ivor Harris becomes CO then SSO at AMI
2013 LT Kewei Zhang arrives at AMI
2014 LTCOL Auliff becomes SSO at AMI, position later upgraded to Administrative Commanding Officer
LT Paul Pickering arrives at AMI Mr Anthony Hunt arrives at AMI to work in animal facility
2015 MAJ Brady McPherson arrives AMI
2016 50th Anniversary of re-establishment of an antimalarial research organisation within the ADF
Highlights
2006
• Transition from Prof Karl H. Rieckmann to new Director Prof G. Dennis Shanks
2007
• Pacific Malaria Initiative (PMI) begun as AusAID program with AMI director on the Malaria Reference Group
as well as Malaria Elimination Group
• Higher (PC3) bio-security arbovirology laboratory completed
• DRD report that relapsing P. vivax infections in ADF personnel returning from East Timor result from clonal
activation of hypnozoites at predetermined intervals
2008
• Port Moresby and Lae, PNG medical screening of PNG Defence Force
• Tanna and Tafea Province, southern Vanuatu islands malaria and mosquito surveys for AusAID PMI
• Santa Cruz and Temotu Province, SI: Malaria and mosquito surveys in isolated southern islands during
RAMSI mission for AusAID PMI
• Aotus colony genetic diversity increased for breeding by importation of remaining monkeys when US Army
discontinued its Aotus program
• Timor-Leste primaquine post exposure prophylaxis trial in 203 ADF soldiers returning from peace-keeping
duties to Australia
• AMI becomes founding member of Asia Pacific Malaria Elimination Network during inauguration meeting in
Brisbane
• DRD’s research on drug interaction genes in P. vivax included in the journal Nature
2009
• Isabel, Solomon Islands malaria survey as part of pre-elimination campaign during RAMSI mission as part
of AusAID funded Pacific Malaria Initiative
• Pandemic influenza studies using records of the First Australian Imperial Force begun with Centre for
Military and Veterans’ Health at the University of Queensland
• Japanese Encephalitis vaccine trial 5 year follow up completed with chimeric Yellow Fever vaccine registered
by the Australian Therapeutic Goods Administration
• Dengue Vaccine trial with Sanofi Aventis conducted at AMI
• DRD report for the first time P. falciparum parasites lacking HRP2 antigen in Peru
• DRD report artemisinin induced dormancy in P. falciparum parasites
• AMI receives its first two NHMRC grants to Drug Resistance and Diagnostics Department
• AMI becomes all officer ADF unit with ending of pathology technician trade
• Liquid Chromatography Mass Spectrometer arrives in Drug Evaluation Department
• Nggella, Solomon Islands malaria and mosquito surveys as part of development of longitudinal malaria
elimination studies (US NIH ICMER)
• Vietnam Australia Defence Malaria Project begins transition with collaboration of Naval Medical Research
Unit 2 from Hawaii / Singapore
• Rotary Against Malaria (RAM) holds its annual meeting at AMI
• Review of AMI’s mission, capabilities and resources for ADF conducted
2013
• External review of Army Malaria Institute recommendations received
• Director AMI joins Bill and Melinda Gates Foundation’s Malaria Advisory Panel
• WHO Collaborating Centre for Malaria renewed
• Malaria entomology work with James Cook University in Solomon Islands History Volume 24 Number 1; January 2016 Page 37
2014
• Vietnam Australia Malaria Research Program continued with support from US Naval Medical Research Unit
– Pacific in Singapore / Cambodia
• Internal review of Army Malaria Institute conducted
• Investigation of post-deployment malaria from OP Olgetta Warrior in PNG
2015
• Visit of RADM Colin Chinn US Pacific Command Surgeon to AMI
• Outbreak of malaria on HMAS Newcastle in Indian Ocean
• Dr Ian Howie-Willis writes monograph Malaria an Unending War on the Australian Army’s long involvement
with malaria

Figure 1 Malaria survey in Isabel, Solomon Islands
 Figure 2 Malaria survey in Tanna, Vanuatu
Figure 2 Malaria survey in Tanna, Vanuatu
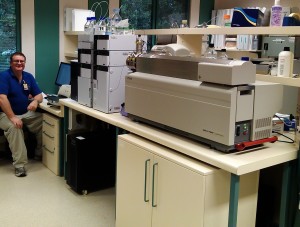
Figure 3 Dengue Vaccine Team conducting Chimerivax
clinical trial

Figure 4 Mass Spectrometer arrives and is installed in
Drug Evaluation Department 2012

Figure 5 Group photo of AMI staff 13 Feb 2014